

Protein display on lactic acid bacteria cell surface
- Tien Giang University, Vietnam, Laboratory of Biosensors, Faculty of Biology - Biotechnology, University of Science, Ho Chi Minh City, Vietnam
- Department of Molecular and Enviromental Biotechnology, Faculty of Biology - Biotechnology, University of Science, Ho Chi Minh City, Vietnam
- Vietnam National University Ho Chi Minh City, Vietnam
- Faculty of Biology - Biotechnology, University of Science, Ho Chi Minh City, Vietnam
Abstract
The strong development of novel protein expression systems, especially in lactic acid bacteria, has attracted a lot of attention and presented many challenges for scientists, not only in terms of safety but also in the ability to preserve food. With the advantage of possessing a rigid cell wall structure and typical acid resistance, lactic acid bacteria have become a potential target not only as a cell factory for producing recombinant proteins used in the food industry but also as a means of delivering antigens for mucosal vaccine development. To achieve this goal, a series of target protein expression systems have been developed in many lactic acid bacteria. This review summarizes recent developments in foreign protein expression systems in both secretion and surface display forms in lactic acid bacteria, including prominent examples such as Lactococcus lactis and Lactobacillus plantarum, as well as current research on developing promising promoter systems to not only improve their ability but also increase their expression efficiency on the surface of lactic acid bacteria.
GIỚI THIỆU
Vi khuẩn lactic acid (lactic acid bacteria, LAB) thuộc nhóm vi khuẩn Gram dương được công nhận an toàn GRAS (Generally Recognized As Safe), thường được ứng dụng trong bảo quản thực phẩm, đã và đang thu hút sự chú ý trong quá trình nghiên cứu và phát triển hệ thống biển hiện protein an toàn cấp thực phẩm (food-grade expression system) 1. LAB thường được tìm thấy trong nhiều dạng môi trường sống tự nhiên như khoang miệng, đường sinh dục, đường tiêu hóa của người và động vật, hoặc sản phẩm lên men của sữa, thịt và rau củ2 và đã được sử dụng suốt một thời gian dài trong quá trình chế biến các loại thực phẩm. LAB bao gồm một số loài trong các chi Lactococcus, Lactobacillus, Enterococcus,và Streptococcus có thể phát triển trong điều kiện kỵ khí lẫn hiếu khí và lên men đường hexose để sản xuất lactic acid như sản phẩm chính cuối cùng3. Quá trình acid hóa khi sản xuất lactic acid là lý do chính cho tác dụng bảo quản của LAB nhằm ức chế sự phát triển của vi sinh vật thường gây hư hỏng thực phẩm4. LAB cũng có thể sản xuất ra các bacteriocin, là các peptide kháng khuẩn, có khả năng ức chế sự phát triển của mầm bệnh cạnh tranh5, 6. Nhiều LAB như Lactobacillus spp., Enterococcus spp., Bifidobacterium spp. đã được công nhận là vi sinh vật có khả năng tăng cường sức khỏe cho đường tiêu hóa của vật chủ, hay còn được gọi là lợi khuẩn. Một ứng dụng tốt khác của lợi khuẩn LAB là kích thích hệ thống miễn dịch của vật chủ. Cụ thể, L. casei Shirota đã cải thiện tình trạng viêm đại tràng ở chuột bằng cách ức chế chuyển vị của NF-κB và sản xuất IL-6 7. Thành phần chưa được xác định của vi khuẩn L. paracasei Ncc2461 đã cảm ứng các tế bào T CD4 trong lách chuột, tạo ra IL-10 và TGF-β 8. Gần đây, Florin Barla và cộng sự9 đã ghi nhận việc sử dụng LAB từ thực phẩm lên men truyền thống của tỉnh Ishikawa để sản xuất γ-aminobutyric acid (GABA) có tác dụng hạ huyết áp. Ngoài ra, LAB được chứng minh giúp giảm cholesterol và hydrolase muối mật 10.
Với những lợi ích trên, LAB là đối tượng tiềm năng trong các ứng dụng liên quan đến thực phẩm chức năng để cộng sinh/cạnh tranh với một số loài vi khuẩn và nấm khác như Escherichia coli, Bacillus subtilis, B. licheniformis, Aspergillus oryzae hoặc A. niger dùng để sản xuất các sản phẩm tự nhiên và tái tổ hợp11. Ngoài ra, nhờ đặc tính cấu trúc cứng của thành tế bào và khả năng kháng acid cao của vi khuẩn Gram dương, nên LAB có tiềm năng được ứng dụng làm xúc tác sinh học toàn tế bào và làm phương tiện vận chuyển kháng nguyên để phát triển vaccine đường niêm mạc. Với mục đích này, một loạt các gene được biểu hiện như là hệ thống protein nhắm trúng đích đã được phát triển trong nhiều LAB12.
Các thuộc tính của những hệ thống biểu hiện này và các ứng dụng hiện tại hoặc tiềm năng đã và đang được nhắm đến chủ yếu ở Lactococcus lactis và Lactobacillus plantarum, là hai loài LAB được nghiên cứu nhiều nhất cho các ứng dụng này. Phần tổng quan này có thảo luận về cấu trúc, chức năng và tình hình nghiên cứu cho đến thời điểm hiện tại của các hệ thống tiết protein qua màng và biểu hiện protein ngoại lai ở vi khuẩn Gram dương nói chung và LAB nói riêng.
Một số protein đã được biểu hiện trên bề mặt tế bào LAB
|
Tên protein (tài liệu tham khảo) |
Chủng chủ |
Chức năng |
Ứng dụng |
|
Surface layer protein SlpA |
L. |
Bám dính |
Liên kết bề mặt |
|
β-mannanase ManB |
L. |
Enzyme |
Chất xúc tác |
|
ϕg1e Lys |
L. |
Phage |
Ly giải các thể thực thuẩn và vi sinh vật gây bệnh |
|
Brevicin 174A BreD, BreG |
L. |
Điều hoà |
Sản xuất bacteriocin (chất kháng khuẩn) |
|
Lam29 |
L. |
Vận chuyển |
Protein liên kết với cysteine của vận chuyển cassette liên kết ATP (ABC) |
CƠ CHẾ TIẾT PROTEIN QUA MÀNG TẾ BÀO CHẤT Ở LAB
Các hệ thống tiết và đích đến cuối cùng của các protein ở vi khuẩn Gram dương
Protein tái tổ hợp có thể được sản xuất trong tế bào chất hoặc được vận chuyển qua màng và xuất trực tiếp ra môi trường, hoặc được đính trên bề mặt vi khuẩn3. Một số cơ chế khác nhau để vận chuyển protein đã được ghi nhận. Ở vi khuẩn Gram dương, bảy cơ chế chính để tiết protein đã được mô tả như Twin-arginine translocation (Tat pathway), Flagella export apparatus (FEA pathway), WXF100 secretion system (Wss pathway), Fimbrillin-protein exporter (FPE pathway), Peptide efflux ABC-transporters, Holin-system, và tiết thông qua con đường Sec (Sec pathway) (Figure 1) 2. Trong đó, protein được tiết ra có thể: (i) Được neo vào màng tế bào (lipoprotein); (ii) Được gắn vào vách tế bào (cell wall/CW) hoặc bằng liên kết cộng hóa trị (protein LPxTG) hoặc không cộng hóa trị (bằng cách biển hiện các motif LysM, GW, CWBD2, SLHD hoặc WXL); (iii) Là một phần của các cấu trúc phụ trên bề mặt tế bào như cellulosome hoặc lông nhung (các tiểu đơn vị được tiết ra qua Sec), pseudo-pili (được lắp ráp thông qua FPE) hoặc tiên mao (được lắp ráp thông qua FEA); (iv) Được tiết ra ngoại bào thông qua Sec, Tat, holin hoặc Wss; hoặc (v) Được chuyển vào tế bào chủ sau khi tiết thông qua Sec của cytolysin phụ thuộc vào cholesterol, được sáp nhập vào màng plasmatic của tế bào chủ và sau đó cho phép chuyển các chất tiết ra bằng quá trình dịch mã qua trung gian cytolysin (CMT).

Các hệ thống tiết và đích đến cuối cùng của các protein được tiết ra ở vi khuẩn Gram dương
Gần đây, con đường Tat ở vi khuẩn và vi khuẩn cổ được chứng minh là con đường dùng để vận chuyển các protein ở dạng gấp cuộn hoàn toàn với một motif twin-arginine được bảo tồn cao có chứa peptide tín hiệu19. Con đường FEA vốn có liên quan đến sự chuyển vị của các thành phần trong tế bào vi khuẩn Gram dương, nhưng cũng được sử dụng để tiết các yếu tố độc lực ra ngoại bào ở vi khuẩn Gram dương20. Một hệ thống tiết chuyên biệt khác được xác định ở vi khuẩn Gram dương là con đường Wss, liên quan đến sự vận chuyển của họ protein WXG100 (ESAT-6)21. Con đường FPE có vai trò trong sự chuyển vị của các tiền chất liên quan đến sự phát triển của vi khuẩn khi cho phép nhập bào DNA ngoại lai qua màng tế bào 18. Holin là các protein sáp nhập xuyên màng nhỏ tạo thành một hệ thống tiết chuyên biệt chỉ có ở vi khuẩn Gram dương 21. Hệ thống holin có liên quan đến hoạt động tiết của hydrolase murein vốn thiếu peptide tín hiệu và đóng vai trò quan trọng trong quá trình tự phân giải. Con đường Sec là con đường tiết protein chủ yếu ở vi khuẩn Gram dương, sử dụng các peptide tín hiệu ở đầu N với các tính năng được bảo tồn ở vi khuẩn Gram dương. Nhờ công cụ tìm kiếm trình tự tương đồng và domain của protein, bộ gene của Lactobacillus đã được chứng minh có chứa các gene mã hóa cho các hệ thống Sec, FPE, holin và không mã hóa cho các yếu tố quan trọng có liên quan đến con đường tiết protein Tat, FEA và Wss 2.
Vai trò của peptide tín hiệu trong cơ chế tiết protein qua màng ở LAB
Peptide tín hiệu (signal peptide, SP), có vai trò trong sự vận chuyển qua màng tế bào chất ở phần lớn các protein tiết, có cấu trúc gồm 3 domain riêng biệt là N, H và C, đóng vai trò quan trọng trong quá trình chuyển vị và tiết của protein22. Một arginine hoặc lysine bảo tồn có trong domain ở đầu N được sử dụng để tương tác với động cơ chuyển vị và phospholipid tích điện âm trong màng lipid kép của màng. Vùng kỵ nước bảo tồn cũng như glycine hoặc proline bảo tồn ở giữa lõi kỵ nước của vùng H (vùng phá xoắn bảo tồn) có thể tạo thành cấu trúc giống kẹp tóc hỗ trợ cho sự chèn vào màng. Vùng phá xoắn bảo tồn có thể được tách ra nhờ vai trò của peptidase tín hiệu (signal peptidase, SPase): vị trí phân tách trên domain ở đầu C của SP được nhận diện bởi một SPase đặc hiệu giúp cắt SP ra khỏi phần còn lại, trở thành protein tiết được vận chuyển qua màng tế bào chất. Mặc dù các SP có cấu trúc tương tự nhau, nhưng các biến thể nhỏ ở các SP khác nhau quyết định đích vận chuyển đến các điểm đến khác nhau hoặc xuất qua các con đường khác nhau 23. Dựa trên các phân tích trình tự của SP và protein nhận biết của chúng, bốn đặc tính đã được chứng minh có mối tương quan với chức năng của SP là tính kỵ nước cao, sự hiện diện của chuỗi xoắn xuyên màng, sự vắng mặt của một motif neo trong protein nhận biết và độ dài của domain H+C24. Những thay đổi về tính kỵ nước của vùng lõi của SP và chiều dài của SP đã được chứng minh là ảnh hưởng đến hiệu quả tiết ở vi khuẩn Gram dương 25. Ngoài sự phụ thuộc cao vào SP, hiệu quả tiết được phát hiện có tính đặc hiệu cao với vật chủ và phụ thuộc vào tính chất của protein mục tiêu như kích thước hoặc mức độ biểu hiện nội bào của chúng24, 25, 26, 27, 28. L. lactis đã được chứng minh có thể tiết ra các protein từ 10 đến 160 kDa thông qua con đường phụ thuộc Sec29. Mức sản xuất cao của protein mục tiêu có thể gây ra tình trạng quá tải động cơ chuyển vị, từ đó dẫn đến hiệu quả tiết thấp 24. Do đó, sự kết hợp tối ưu giữa SP và protein mục tiêu mong muốn là điều cần thiết và thiết yếu để có thể đạt được hiệu quả tiết cao. Một số vi khuẩn Gram dương có khả năng tiết ra protein với hiệu suất khá cao. Các mô hình trên vi khuẩn Gram dương như B. subtilis, L. lactis và L. plantarum đã được nghiên cứu kỹ về tiềm năng dùng trong sản xuất protein ngoại lai ở dạng tiết25, 26.
Usp45 là SP được khai thác rộng rãi nhất ở LAB biến đổi gene ở Lactococcus 28. Các SP dị loài khác đã được sử dụng để tiết các protein ngoại lai trong L. plantarum bao gồm SP từ Streptococcus pyogenes là protein M6, từ L. amylovious là amylase, và từ L. brevis là protein S-layer (SlpA). Geir Mathiesen (2008) đã thử nghiệm các SP tương đồng từ L. plantarum WCFS1 và so sánh chức năng của chúng trong quá trình tiết hai mô hình protein ngoại lai khác nhau (NucA từ Staphylococcus aureus và AmyA từ L. amylovorus) 30. Trong nghiên cứu này, hệ thống pSIP cảm ứng biểu hiện dựa trên pheromone peptide trong Lactobacillus đã được sử dụng31 và hiệu quả tiết của các SP này đã được so sánh với hai SP (ngoại lai) nổi tiếng Usp45 và M6. Kết quả cho thấy 4 trong số 10 trình tự được chọn có vị trí phân cắt là Ala-X-Ala-Ala, nhưng chúng không đạt hiệu quả tiết cao. Hai SP từ L. plantarum là Lp-0373 và Lp-0600, vốn có trình tự motif là Val-X-Ala ở vị trí -3 đến -1 so với vị trí phân cắt, là các SP hoạt động tốt nhất để tiết nuclease và amylase30. Trong một nghiên cứu khác, Karlskes và đồng nghiệp cũng chỉ ra rằng ba SP có nguồn gốc từ Lp-3050, Lp-0373 và Lp-2578 cho thấy mức độ sản xuất và hiệu quả tiết của nuclease NucA ở năm loài Lactobacillus bao gồm L. rhamnosus, L. brevis, L. gasseri, L. curvatus và L. plantarum khá khác nhau; trong đó SP có nguồn gốc từ Lp-3050 cho lượng NucA tiết ra cao nhất 32. Kết quả này cho mỗi sự kết hợp giữa chủng chủ và protein mục tiêu có thể cần phải được tối ưu hóa riêng biệt.
CÁC HỆ THỐNG BIỂU HIỆN PROTEIN TRÊN BỀ MẶT Ở LAB
Việc biểu hiện các protein ngoại lai trên bề mặt tế bào vi khuẩn là một chiến lược hấp dẫn cho nhiều ứng dụng trong lĩnh vực công nghệ sinh học, điều trị bệnh và công nghiệp như phát triển vaccine sống, sản xuất kháng thể, xúc tác sinh học toàn tế bào, hấp thụ sinh học và cảm biến sinh học33. Một số protein đã được biểu hiện trên bề mặt tế bào LAB được trình bày ở Table 1.
Một trong những tính năng hấp dẫn nhất của nghiên cứu biểu hiện trên bề mặt tế bào là việc các protein mục tiêu dung hợp với các motif neo vốn được đồng thời vừa tổng hợp vừa cố định trên bề mặt tế bào vi khuẩn; kết quả là dễ dàng thu nhận được protein mục tiêu từ các tế bào vi khuẩn trong quá trình nuôi cấy. Trong cách tiếp cận này, các tế bào tự thực hiện toàn bộ quy trình phức tạp gồm nhiều phản ứng ở nhiều bước vốn tốn nhiều năng lượng và thời gian, do đó mang lại lợi ích lớn về mặt kinh tế. Ngoài ra, các protein ngoại lai neo trên bề mặt của tế bào vi khuẩn có tính ổn định cũng như khả năng bảo toàn trong điều kiện khắc nghiệt cao hơn nhiều lần so với protein tự do, đặc biệt là khi protein được chèn vào thành của tế bào vi khuẩn 34. Điều này có nghĩa chiến lược biểu hiện protein trên bề mặt cần được cân nhắc, phân tích kỹ lưỡng và lựa chọn hợp lý, đủ cao để cải thiện khả năng tiếp cận của protein với các đích đến của chúng và đủ thấp để bảo vệ các protein được biểu hiện trên bề khỏi bị phân hủy 3. Mặt khác, mức độ biểu hiện của protein phụ thuộc vào nhiều yếu tố bao gồm protein mục tiêu, chủng chủ và các loại neo được sử dụng. Một protein mục tiêu tự nó cũng có ảnh hưởng đến quá trình dịch mã và biểu hiện lên bề mặt. Kích thước quá lớn hoặc mức độ biểu hiện quá cao của protein mục tiêu có thể dẫn đến tiết không hiệu quả, điều này có thể làm giảm sự tăng trưởng của vật chủ tái tổ hợp và lượng thấp protein biểu hiện trên bề mặt của nó 35. Các protein mục tiêu khác nhau được dung hợp vào cùng một loại neo sẽ được chuyển đến các vị trí khác nhau trong cùng một chủng chủ36. Rõ ràng, việc lựa chọn chủng chủ để sử dụng có vai trò cực kỳ quan trọng trong chiến lược biểu hiện trên bề mặt. Một chủng chủ tốt phải phù hợp với “protein đi theo/passenger protein”, dễ dàng nuôi cấy mà không cần ly giải tế bào và hoạt tính protease ngoại bào và protease tương ứng của thành tế bào thấp. Xem xét điểm này, vi khuẩn Gram dương dường như là sự lựa chọn thuận lợi hơn là vi khuẩn Gram âm vì chúng có cấu trúc thành tế bào khá cứng34 và chỉ có một màng kép thuận lợi cho protein đi xuyên qua 35. Các neo chứa một SP hiệu quả để xuất protein mục tiêu qua màng trong. Một cấu trúc neo ổn định cũng như chiều dài và trình tự của neo nên được cân nhắc để có thể giữ được protein dung hợp trên bề mặt tế bào mà không bị tách ra. Ngoài việc tương thích với các chuỗi ngoại lai được dung hợp, neo còn có khả năng chống lại sự tấn công của các protease có mặt trong không gian chu chất hoặc trong môi trường nuôi cấy. Tất cả các lưu ý cần thiết được đề cập ở trên gợi ý rằng sự hiểu biết sâu về chức năng của neo có thể có ích cho việc tối ưu hóa hệ thống biểu hiện trên bề mặt. Về nguyên tắc, có hai cách khác nhau để gắn protein được tiết ra lên trên bề mặt của vi khuẩn là thông qua liên kết cộng hóa trị với màng tế bào hoặc thành tế bào và liên kết không cộng hóa trị thông qua protein domain tương tác mạnh với các thành phần của thành tế bào hoặc màng (Figure 2). Cả hai hệ thống này đã được sử dụng trong LAB, chủ yếu ở L. lactis và các lactobacilli khác nhau.

Các phương pháp dùng để biểu hiện protein lên bề mặt tế bào Lactobacillus
Neo không cộng hoá trị
Neo xuyên màng ở đầu N
Sự tiết protein vào môi trường ngoại bào thông qua con đường Sec phụ thuộc vào SPase tạo điều kiện giải phóng protein trưởng thành bằng cách tách SP ở đầu N từ preprotein (SP+protein tiết)3. Tuy nhiên, ở cơ chế neo xuyên màng ở đầu N, trình tự amino acid trong peptide tín hiệu ở preprotein thiếu vị trí phân cắt của SPase, dẫn đến sự hình thành neo trong đó protein sẽ được gắn vào thành tế bào bởi các liên kết không cộng hóa trị. Trong trường hợp này, SP không được phân cắt với liên kết kỵ nước ở trung tâm sẽ có chức năng như một chuỗi xoắn xuyên màng ở đầu N có thể gắn protein vào màng. Để có thể hình thành được neo xuyên màng đầu N, protein mục tiêu cần phải được dung hợp với chuỗi neo có độ dài thích hợp. Một neo ngắn sẽ có khả năng bảo vệ protein mục tiêu bằng cách chèn hoàn toàn vào thành của tế bào, nhưng hạn chế là thiếu khả năng tiếp cận. Poquet và cộng sự (1998) đã xác định được hai đoạn gene có nguồn gốc từ Lactococcus mã hóa cho các neo có kích thước đa dạng từ 40 đến 234 amino acid dựa trên phương pháp sàng lọc ngẫu nhiên với tín hiệu xuất thu nhận từ thư viện gene của L. lactis và sử dụng nuclease của Staphylococcus aureus làm reporter protein. Một trong những neo xoắn xuyên màng ở đầu N được khai thác nhiều nhất là protein PgsA từ B. subtilis, bao gồm một vùng xuyên màng dài từ 25 đến 44 amino acid được phân tích bởi hệ thống SOSUI. Protein PgsA đã được chứng minh là một phần của phức hợp poly-γ-glutamate synthetase và có chức năng ổn định phức hợp bằng cách neo vào trong màng của tế bào. Narika đã chỉ ra rằng sự dung hợp của α-amylase vào đầu C của PgsA dẫn đến việc biểu hiện thành công trên bề mặt E. coli và L. casei 38. Việc định vị protein dung hợp PgsA-AmyA-FLAG trên bề mặt tế bào sẽ được phát hiện bằng phương pháp flow cytometry và miễn dịch huỳnh quang. Các tế bào L. casei biểu hiện AmyAF trên bề mặt thể hiện hoạt động thủy phân tăng đáng kể so đối với tinh bột dạng hòa tan. Gần đây, một neo xuyên màng từ Lp-1568, đây là một protein neo đầu N và có nguồn gốc từ L. plantarum đã được ứng dụng để biểu hiện các phiên bản dài và ngắn của invasin (Inv và InvS tương ứng) từ tác nhận gây bệnh ở người là Yersinia pseudotuberculosis trên bề mặt của tế bào vi khuẩn L. plantarum. Sự định vị trên bề mặt của cả hai phiên bản invasin đã được xác nhận bằng phương pháp flow cytometry và miễn dịch huỳnh quang, nhưng chỉ có InS được neo bởi Lp-1568 mới có hoạt động NF-kB trong dòng tế bào monocytic 39. Các kết quả được công bố nêu trên cho thấy chiến lược neo xuyên màng ở đầu N-terminal để biểu hiện các protein ngoại lai trên bề mặt tế bào LAB là một phương pháp có nhiều tiềm năng.
Neo gắn miền trung gian
Sự hình thành liên kết không cộng hóa trị trong chiến lược gắn protein ngoại lai vào thành tế bào có thể thu được bằng cách sử dụng các domain gắn thành tế bào khác nhau như surface (S)-layer protein (SLP), domain LysM hoặc domain WxL40, 41, 42. Các S-layer là mạng lưới các đơn phân của tiểu đơn vị protein hoặc glycoprotein với trọng lượng phân tử 40−200 kDa 43. Cấu trúc S-layer gồm hai vùng chức năng là domain tự lắp ráp và domain gắn tế bào, chịu trách nhiệm cho quá trình neo vào bề mặt tế bào thông qua việc hình thành các liên kết không cộng hóa trị với cấu trúc trên bề mặt của tế bào, thường là với lipopolysaccharide (LPS) có sẵn trong vi khuẩn Gram âm và với các polysaccharide có trên thành tế bào ở vi khuẩn Gram dương 42. Những vùng này đã được mô tả rõ ràng trong S-layer protein S của L. acidophilus ATCC 4356 44, SlpA của L. brevis ATCC 828745 và CbsA của L. crispatus JCM 5810 46. Các nghiên cứu đã chứng minh S-layer protein trong một số loài Lactobacillus có trọng lượng phân tử nhỏ (25−71 kDa), giá trị pI cao 9,35−10,4 47 và chiếm 10−15% tổng số protein. So với SLP từ các loài khác sử dụng motif tương đồng lớp S (S-layer homology, SLH) cho sự gắn kết không cộng hóa trị vào thành tế bào, thì SLP từ Lactobacillus đóng vai trò là trung gian cho quá trình gắn kết với thành tế bào thông qua tương tác giữa các amino acid cơ bản trong vùng gắn kết với thành tế bào và các polymer thành tế bào tích điện âm như lipoteichoic acid hoặc teichoic acid 48. Các tính năng khác biệt kể trên của SLP từ Lactobacillus đã mang đến nhiều tiềm năng trong khai thác các cognate promoter và SP có nguồn gốc từ SLP để biểu hiện và tiết các protein ngoại lai trong LAB, cũng như sử dụng SLP làm chất mang cho biểu hiện protein ngoại lai trên bề mặt của Lactobacilus và Lactococcus 48, 49, 50. Một số SLP trong các loài Lactobacillus đã được nghiên cứu rộng rãi cho các mục đích khác nhau như: phương tiện để vận chuyển kháng nguyên, hoặc chất xúc tác sinh học toàn bộ tế bào 51. S-layer subunit (SlpA) của L. brevis đã biểu hiện thành công mô hình epitope ngoại lai trên bề mặt tế bào trong khi vẫn duy trì được cấu trúc mạng tinh thể của S-layer 52. Protein gắn collagen (collagen-binding protein, Cnb) từ L. reuteri NCIB 11951, vốn có quan hệ tương đồng khá gần với neo S-layer là BspA, đã được sử dụng làm protein neo để biển hiện β-glucanase trên bề mặt tế bào của L. reuteri Pg4. Vị trí trên bề mặt và khả năng phân hủyβ-glucan của β-glucanase biểu hiện trên bề mặt đã được công nhận 53. Một nghiên cứu khác đã chỉ ra rằng SLP từ L. crispatus K2-4-3 được phân lập từ ruột gà có thể được sử dụng để biểu hiện các protein ngoại lai trên bề mặt của một số chủng Lactobacillus không có S-layer thí dụ như L. lactis, suy ra tiềm năng sử dụng neo SLP để biểu hiện trên bề mặt của bản thân vật chủ vốn không thể tự sản xuất ra SLP54.
Một phương pháp neo khác dựa vào liên kết không cộng hóa trị giữa protein với thành của tế bào là thông qua domain LysM. Đây là một module phổ biến trên thành tế bào, có khả năng phân hủy enzyme và protein, là tác nhân gây độc lực ở vi khuẩn 55. Các domain LysM với độ dài khác nhau từ 44 đến 65 amino acid, tồn tại một hoặc nhiều bản sao trong mỗi protein 3, thường hiện diện ở vùng đầu N hoặc đầu C của protein và gắn kết với peptidoglycan và chitin56. Khả năng gắn kết này của các domain LysM đã được áp dụng rộng rãi cho các chiến lược biểu hiện trên bề mặt không biến đổi gene dựa trên việc tạo ra protein dung hợp có chứa LysM trong một chủng chủ sản xuất, ví dụ như E. coli hoặc L. lactis, sau đó là cố định lên trên bề mặt một chủng chủ không tái tổ hợp nào đó ví dụ như LAB 57. Với phương pháp này, chủng chủ có thể được trộn lẫn với protein dung hợp đã hoặc chưa qua tinh sạch, và lượng protein được nạp lên trên bề mặt tế bào có thể được kiểm soát dễ dàng hơn khi được so sánh với cách biểu hiện tái tổ hợp thông thường3. Nhiều protein neo có chứa motif LysM đã được phát hiện và ứng dụng cho mục đích này57, 58. Neo protein (protein anchor, PA) AcmA có nguồn gốc từ peptidoglycan hydrolase của L. lactis là một ví dụ. Neo này chứa ba motif LysM dài 45 amino acid được phân tách bằng các spacer sequences. Bosma và cộng sự đã sử dụng PA để biểu hiện hoặc α-amylase (α-PA) hoặc β-lactamase (β-PA) trên bề mặt của các tế bào lactococcus đã qua xử lý bằng acid. Cả hai enzyme đều hoạt động khi chúng được neo một cách vô tình bởi PA. Raha và cộng sự (2005) cũng đã sử dụng chuỗi lặp lại 255 bp của vùng gắn kết với thành tế bào của protein AcmA đã được đề cập ở trên để biểu hiện vùng A1 và A3 của protein VP1 của enterovirus 71 (EV71) trên bề mặt. Việc biểu hiện thành công các epitope VP1 của EV71 trên bề mặt của L. lactis được xác nhận bằng whole-cell ELISA đã mở ra hướng mới để phát triển vaccine uống. Một ví dụ khác liên quan đến chiến lược không biến đổi gene này là MurO từ L. plantarum hoặc Ly5C từ L. fermentum khi biểu hiện thành công β-galactosidase trên bề mặt L. plantarum59, hoặc trên các tế bào LAB 58 khi ứng dụng trên phạm vi rộng hơn. Các nghiên cứu này cung cấp phương thức gắn protein ngoại lai trên bề mặt của các chủng lợi khuẩn LAB không sử dụng phương pháp biến đổi gene.
Neo cộng hoá trị
Neo lipoprotein
Liên kết cộng hóa trị với màng tế bào có thể được hình thành qua các neo lipoprotein, thường bao gồm một SP ở đầu N có chứa motif lipobox trong vùng đầu C. Sau khi tiết ra protein theo con đường Sec, một diacylglycerol transferase xúc tác việc gắn cộng hóa trị một cysteine bảo tồn trong lipobox với một phospholipid trong lớp màng, trong khi SP được loại bỏ bởi peptidase đặc hiệu của lipobox. Do đó, việc neo protein tái tổ hợp vào màng tế bào có thể đạt được bằng cách dung hợp một protein ngoại lai ở vùng hạ lưu của một neo lipoprotein thích hợp. Việc sử dụng neo lipoprotein để biểu hiện protein ngoại lai trong LAB đã nhận được tương đối ít sự chú ý so với các phương pháp biểu hiện trên bề mặt khác. Trong L. plantarum, hai neo lipoprotein khác nhau là Lp-1261 và Lp-1452 có nguồn gốc tương ứng từ oligopeptide ABC transporter (Lp-1261) và peptidylprolyl isomerase PrsA (Lp-1452), đã được sử dụng để biểu hiện trên bề mặt một đoại dài (Inv) và một đoạn ngắn (InvS) của extracellular domain của invasin từ tác nhân gây bệnh Yersinia pseudotuberculosis trên người. SP và vùng của domain xúc tác chịu trách nhiệm cho quá trình neo cộng hóa trị vào lớp ngoài của màng plasma thông qua cysteine ở đầu N đã bị chuyển hóa lipid. Sự biểu hiện thành công lên bề mặt của Inv và InvS trên bề mặt tế bào Lactobacillus của cả hai neo Lp-1261 và Lp-1452 đã được xác nhận bằng phân tích miễn dịch huỳnh quang và flow cytometry. Đáng chú ý, cả hai neo lipoprotein cho kết quả hoạt động tốt nhất trong số các neo đã được thử nghiệm vốn chứa invasin nguyên bản có cấu trúc khá lớn và gây ra phản ứng viêm mạnh mẽ 39. Trong một nghiên cứu khác, Kuczkowska và cộng sự 60 cũng đã sử dụng hai neo Lp-1261 và Lp-1452 để gắn CCL3Gag, là loại protein dung hợp bao gồm kháng nguyên HIV-1 Gag bị cắt ngắn và chemokine CCL3 của chuột, lên trên bề mặt của L. plantarum NC8. Kết quả nghiên cứu cho thấy không thể xác định vị trí gắn trên bề mặt của protein neo Lp-1261 khi quan sát bằng kính hiển vi, có thể là do quá trình tiết protein hiệu quả thấp, có quá nhiều vị trí chèn vô thành của tế bào. Trong khi đó, các tế bào vi khuẩn biểu hiện CCL3Gag bằng neo Lp-1452 cho thấy tín hiệu huỳnh quang mạnh hơn nhiều, có thể do Lp-1452 giúp cho domain CCL3 tiếp xúc nhiều hơn với kháng thể.
Neo LPxTG
Một neo khác, trong đó có sự hình thành liên kết cộng hóa trị với thành tế bào, có thể được tạo ra bằng cách sử dụng con đường sortase (SrtA) 61. Cụ thể, protein tiết ra mang một neo ở đầu C, mà bản thân neo này có chứa motif LPxTG (LPQTxE trong L. plantarum), theo sau là một domain kỵ nước và một đuôi tích điện dương. Domain kỵ nước và đuôi tích điện giữ cho protein không bị giải phóng vào môi trường do đó cho phép nhận ra motif LPxTG bởi một transpeptidase liên quan đến màng gọi là sortase33. Sortase xúc tác phản ứng cắt liên kết peptide, thí dụ như liên kết giữa threonine và glycine trong motif LPxTG và liên kết giữa threonine ở đầu C của protein bề mặt với cầu nối pentapeptide của peptidoglycan 62. Phản ứng đặc hiệu này đã được khai thác hàng loạt trong lĩnh vực công nghệ sinh học như tổng hợp nên các liên kết nucleic acid-peptide và neoglycoconjugate, tuần hoàn protein, đánh dấu protein bề mặt tế bào trên tế bào sống, phương tiện phân phối vaccine sống và xúc tác sinh học toàn tế bào63. Piard và các đồng nghiệp đã kết luận rằng hiệu quả neo của protein M6 từ Streptococcus pyogenes với thành tế bào cho kết quả rất khác biệt trong các chủng chủ khác nhau bao gồm L. lactis, L. fermentum, L. sake và S. thermophilus. Dieye và cộng sự28 nghiên cứu về biểu hiện thành công nucease NucA trên bề mặt của tế bào vi khuẩn L. lactis bằng cách sử dụng tín hiệu phân loại (sorting signal) ở đầu C của protein M6. Các tác giả này đã có một đánh giá tổng quan về trọng lượng phân tử của các protein dung hợp có chứa neo với thành tế bào của protein M6. Các dải có trọng lượng phân tử lớn đã được phát hiện trong mảnh vỡ thành tế bào cho thấy sự hiện diện của các mảnh peptidoglycan có kích thước khác nhau được liên kết cộng hóa trị với protein bề mặt. Turner và các đồng nghiệp 64 đồng ý với quan sát trên khi protein dung hợp Mlp-His6 chứa tín hiệu neo với thành tế bào là LPxTG được phát hiện bằng kỹ thuật Western blot với khối lượng phân tử cao hơn. Các tác giả phỏng đoán kết quả trên có được là do quá trình glycosyl hóa protein này. Gần đây, một neo vách tế bào mới từ L. plantarum WCFS1 đã được tìm thấy và sử dụng trong biểu hiện 37-kDa oncofetal antigene (OFA) trên bề mặt tế bào L. plantarum WCFS165. Dựa trên trình tự của Lp-2578, là một protein được ghi nhận là protein “bám dính” trên bề mặt của thành tế bào từ L. plantarum WCFS1, Fredriksen và các cộng sự đã thiết kế ba neo chứa LPxTG với độ dài khác nhau tương ứng với vùng có độ phức tạp thấp (low-complexity region) đã bị cắt ngắn, vùng có độ phức tạp thấp còn nguyên vẹn và gần như là protein hoàn chỉnh. Dữ liệu từ nghiên cứu đã cho thấy rằng sự khác biệt về chiều dài của neo ảnh hưởng đến hiệu quả của quá trình neo và độ ổn định của protein; tuy nhiên sự khác biệt này không lớn cũng như không rõ ràng để dự đoán.
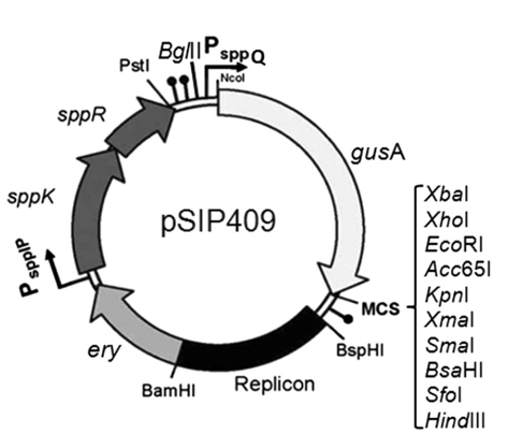
Đáng chú ý là trong một nghiên cứu gần đây của Hoàng Minh và cộng sự 14, hệ thống vector pSIP đã được sử dụng để nghiên cứu chức năng của hai phương pháp gắn cộng hóa trị để biểu hiện các enzyme liên quan đến lĩnh vực thực phẩm như mannanase hoặc chitosanase trên bề mặt tế bào của L. plantarum như là chất xúc tác sinh học toàn tế bào (Figure 3). Hai khác biệt chính khi so sánh hai phương pháp này là: (i) Protein được gắn vào màng hoặc thành tế bào, kết quả là phương pháp neo vào thành tế bào qua trung gian sortase sẽ biểu hiện nhiều protein ngoại vi hơn phương pháp còn lại; (ii) Đầu N của protein được gắn vào bề mặt của tế bào trong phương pháp sử dụng lipobox, còn phương pháp neo vào vách tế bào qua trung gian sortase lại gắn protein ở đầu C. Việc sử dụng hai phương pháp neo cộng hóa trị này để biểu hiện các protein mục tiêu khác nhau trên bề mặt tế bào Lactobacillus đã cho cái nhìn sâu sắc và toàn diện về hệ thống neo của vi khuẩn.
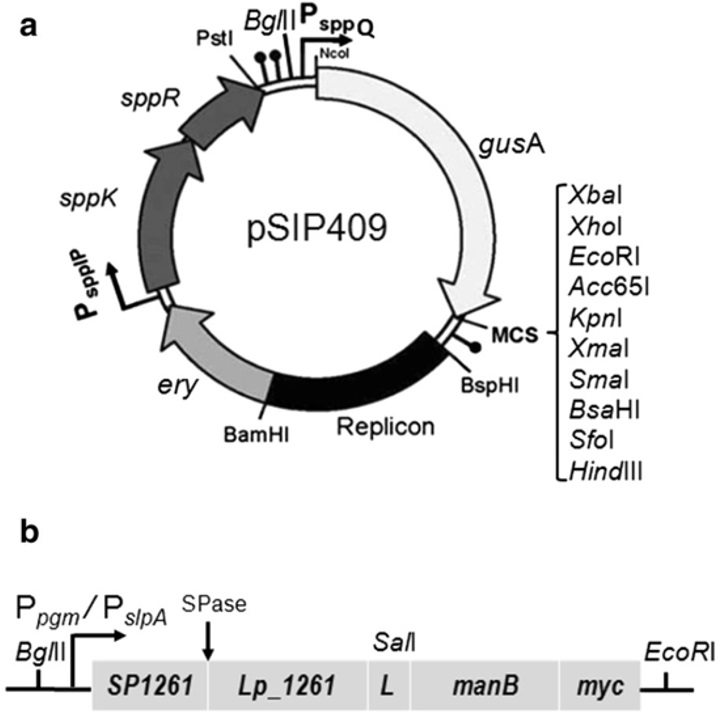
Hệ thống vector biểu hiện trên bề mặt tế bào LAB. (a) Cấu trúc vector biểu hiện pSIP409 sử dụng motif neo lipoprotein đầu N của mannanase (ManB) trong
PHÁT TRIỂN CÁC PROMOTOR BIỂU HIỆN THƯỜNG TRỰC Ở LAB
Như đã đề cập ở trên, việc phát triển các công cụ di truyền để sản xuất protein và enzyme ngoại lai trong LAB đang rất được quan tâm. Một số các hệ thống vector biểu hiện cảm ứng đã được phát triển trong LAB. Các vector pSIP dựa trên promoter cảm ứng mạnh P vốn dành riêng cho chủng chủ Lactobacillus, đã được chứng minh là cung cấp mức độ biểu hiện cao, kiểm soát chặt chẽ bằng lượng nano gram được thêm vào của tác nhân cảm ứng là peptide cảm ứng (induction peptide, IP). Nhìn chung, hệ thống vector biểu hiện cảm ứng hữu ích trong trường hợp cần phải đạt được mức sản xuất tối đa của protein tái tổ hợp tại một thời điểm nhất định và cụ thể của quá trình nuôi cấy, hoặc khi protein có tác động tiêu cực đến quá trình chuyển hóa tế bào vi khuẩn66. Ngoài ra, ngày càng có nhiều sự quan tâm đối với sự phát triển của LAB với vai trò là nhà máy sản xuất tại chỗ hoặc là phương tiện phân phối các hợp chất điều trị hoặc enzyme trong cơ thể người, nhưng điều này khiến cho hệ thống biểu hiện cảm ứng không còn phù hợp 67. Đối với các ứng dụng này, chiến lược sử dụng các vector biểu hiện thường trực liên quan đến sản xuất protein ở mức mong muốn sẽ là một giải pháp thay thế khả thi. Đáng chú ý là các promoter vi khuẩn có chung đặc tính có thể có tác động và cường độ khác nhau trong LAB; do đó, cần thiết phải xác định, đánh giá các promoter và kiểm tra tính hiệu quả của các hệ thống vector biểu hiện khác nhau cho mỗi vật chủ mới68. Nhìn chung, có ba chiến lược chính để tìm kiếm và xác định các promoter có nguồn gốc từ L. lactis và các LAB khác hiện hữu bao gồm: (a) Sử dụng các vector sàng lọc mang promoterless reporter genes; (b) Phân lập các promoter từ các gene house-keeping mạnh và thường trực; (3) Tạo ra các promoter tổng hợp từ các consensus promoter 12. Rud và cộng sự đã phát triển một thư viện promoter tổng hợp (synthetic promoter library, SPL) cho L. plantarum dựa trên cách tiếp cận của Jensen & Hammer, trong đó một thư viện các promoter nhân tạo cho L. lactis thu được bằng cách sắp xếp ngẫu nhiên spacer sequence vốn phân tách các consensus sequences của promoter. SPL thu được đã cung cấp một loạt các promoter thường trực sở hữu các thế mạnh khác nhau trong sản xuất ổn định protein; và các promoter mạnh nhất, điển hình là P, đã mang lại sản lượng protein tương đương với native rRNA promoters 67. Tauer và đồng nghiệp 66 đã so sánh chức năng của bốn promoter thường trực bao gồm P, P, P và P và kiểm tra biểu hiện trong tế bào chất của gene mCherry với mục đích cung cấp một hệ thống thường trực phù hợp cho các ứng dụng trong lên men thực phẩm hoặc phân phối thuốc trong điều kiện in vivo. Promoter P được đánh giá là promoter mạnh nhất. Việc biểu hiện cũng đạt được kết quả tốt khi sử dụng cả hai promoter mới là P và P, hiện có sẵn dưới dạng các yếu tố bổ sung để thúc đẩy quá trình biểu hiện thường trực trong L. plantarum và L. buchneri trong khi promoter P cho hiệu quả rất yếu. Do đó, việc đánh giá độ mạnh của các promoter thường trực này có thể có ích cho sự phát triển các hệ thống biểu hiện thường trực trong LAB, từ mức độ sản xuất trung bình cho đến mạnh.
Trong một nghiên cứu gần đây, ảnh hưởng của ba promoter thường trực là pgm, slpA và slpB đến biểu hiện dạng tiết và biểu hiện trên bề mặt của gene manB trong chủng chủ L. plantarum đã được tiến hành so sánh. Những promoter thường trực này được đánh giá là yếu tố tiềm năng cho khả năng biểu hiện trong nội bào của các protein ngoại lai trong lactobacilli. Promoter cho đột biến phosphoglycerate (phosphoglycerate mutase, pgm) là một promoter thường trực mạnh có nguồn gốc từ L. acidophilus NCFM, vốn là một chủng lợi khuẩn đã có lịch sử được sử dụng rộng rãi trong các sản phẩm bổ sung dinh dưỡng, các sản phẩm từ sữa và sữa công thức cho trẻ sơ sinh (Figure 4) 66, 47. Promoter pgm được xác định dựa trên phân tích bộ gene đã chứng minh một số operon biểu hiện khác biệt để đáp ứng với các nguồn carbohydrate hoặc biểu hiện thường trực bất kể nguồn carbohydrate 69. Tri Duong và cộng sự đã phát hiện ra mức độ biểu hiện của GUS, vốn chịu kiểm soát bởi promoter thường trực pgm, cao xấp xỉ 10 lần so với mức biểu hiện trong cấu trúc cảm ứng ở hai chủng chủ là L. acidophilus và L. gasseri. Promoter đã có thể điều khiển biểu hiện của operon oxalate, cải thiện đáng kể quá trình phân hủy oxalate của L. gasseri ATCC 33323. Kết quả này cho thấy việc sử dụng một hệ thống vector biểu hiện thường trực dựa trên promoter pgm có thể là một công cụ hữu ích để biểu hiện vượt mức các protein và enzyme mong muốn trong lactobacilli và các chủng khác của LAB.

Vùng trình tự gốc của promoter
Promoter slpA của L. acidophilus ATCC 4356 đã được nghiên cứu rộng rãi trong phát triển các hệ thống biểu hiện gene ngoại lai trong một loạt các chủng chủ Lactobacillus. Promoter slpA đã được chứng minh là có hiệu quả cao không chỉ trong chủng chủ tự nhiên mà còn trong cả chủng L. casei ATCC 393, vốn không sở hữu S-layer protein 68, 48. Đáng chú ý, vùng promoter của vị trí biểu hiện gene slpA từ L. acidophilus bao gồm hai promoter tiềm năng là P-1 (vị trí -228 đến -198) và P-2 (vị trí -335 đến -303) (Figure 5). Kết quả này cho thấy lượng mRNA được định hướng bởi một cấu trúc nhiều promoter có thể cao hơn so với khi chỉ được định hướng của một promoter 48.

Trình tự nucleotide trước khi bắt đầu dịch mã gene
KẾT LUẬN
Bài báo này trình bày tổng quan về các hệ thống biểu hiện protein ngoại lai ở dạng tiết và biểu hiện trên bề mặt của LAB, và một số giải pháp nhằm tăng hiệu quả biểu hiện của chúng. Mặc dù có rất nhiều tiềm năng đã và đang được chứng minh, nhưng thực tế là để có thể áp dụng phổ biến các hệ thống biểu hiện ở LAB, các nhà khoa học cần phải có thêm nhiều nghiên cứu sâu hơn với hy vọng rằng trong tương lai, các nghiên cứu này sẽ có thêm nhiều đột phá nhằm đưa ứng dụng của các chúng đến tất cả các lĩnh vực của cuộc sống con người.
DANH MỤC CÁC TỪ VIẾT TẮT
LAB: Lactic Acid Bacteria
GABA: Gama-Aminobutyric Acid
CM: Cell Membrane
Cyto: Cytoblast
CW: Cell Wall
OM: Double-Stranded Ribonucleic Acid
SRP: Signal Recognition Particle
SP: Signal Peptide
SPase: Signal Peptidase
SLP: Surface (S)-Layer Protein
SLH: S-Layer Homology
PA: Protein Anchor
LPS: Lipopolysaccharide
NICE: Nisin Controlled Gene Expression System
SICE: Stress - Induced Controlled Expression
IP: Induction Peptide
HK: Histidine Kinase
SPL: Synthetic Promoter Library
LỜI CẢM ƠN
Nghiên cứu được tài trợ bởi Sở Khoa học Công nghệ tỉnh Tiền Giang trong khuôn khổ Đề tài mã số ĐTNN 05/19.
Nguyễn Thanh Tấn được tài trợ bởi Chương trình học bổng đào tạo thạc sĩ, tiến sĩ trong nước của Quỹ Đổi mới sáng tạo Vingroup (VINIF), mã số VINIF.2022.ThS.077.
XUNG ĐỘT LỢI ÍCH
Các tác giả cam kết không có xung đột lợi ích.
ĐÓNG GÓP CỦA CÁC TÁC GIẢ
Các tác giả Nguyễn Thị Phương Thảo, Nguyễn Thanh Tấn viết, tổng hợp, và chỉnh sửa bản thảo. Tác giả Trần Văn Hiếu tham gia chỉnh sửa bản thảo. Tất cả các tác giả đồng ý với bản thảo cuối cùng khi nộp.